

お知らせ
研究ブログ
-
坂元七望(B4)が、卒業論文で優秀発表賞(工学部応用化学科)を受賞しました。(2024.2.9)
2024.02.13 受賞
-
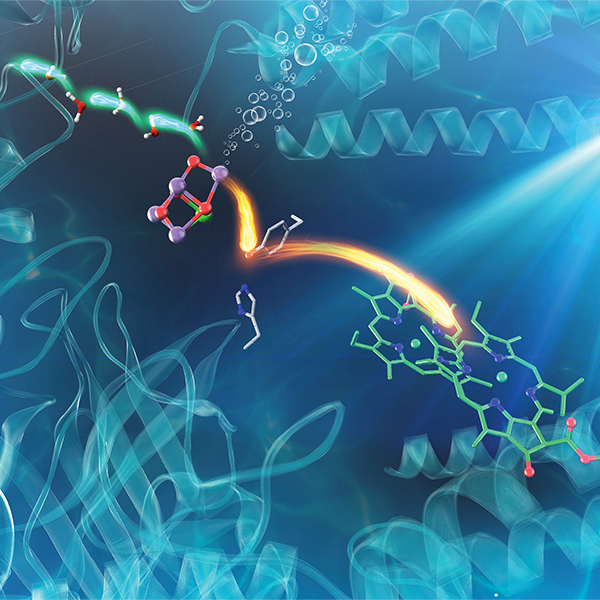
プレスリリース「光合成反応における光損傷と修復のメカニズム解明 ―傷ついたタンパク質を見つけて分解するしくみを明らかに―」(2023.11.21)
2023.11.21 プレスリリース
-
西尾俊哉(M2)が、修士論文で藤嶋賞(工学系研究科応用化学専攻)を受賞しました。(2023.2.1)
2023.02.02 受賞
-
石北 央(教授)が米化学会 Biochemistry 誌のEditorial Advisory Board に加わりました。
2023.01.10
-
光合成という夢のある反応 (西尾 俊哉 [M2])
2022.11.11 エッセイ -
プレスリリース「植物の光合成初期過程の酸素発生活性を向上させるアミノ酸変異を発見—光合成・人工光合成の光エネルギー変換効率の向上へ期待—」(2022.8.18)
2022.08.18 プレスリリース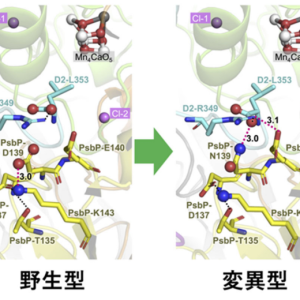
-
学生インタビュー[菅生 優 (M2), 千葉 義大 (M1)]
2022.09.14 石北研についてインタビュー
-
理論によって放たれる光 (神田 知樹 (M2))
2022.06.10 エッセイ
-
坂元七望(B4)が、卒業論文で優秀発表賞(工学部応用化学科)を受賞しました。(2024.2.9)
2024.02.13 受賞 -
西尾俊哉(M2)が、修士論文で藤嶋賞(工学系研究科応用化学専攻)を受賞しました。(2023.2.1)
2023.02.02 受賞 -
高校生・受験生が東大をもっと知るためのサイト 「キミの東大」に辻村真樹(D1)が掲載されました。(2022.5.31)
2022.06.10 辻村真樹受賞お知らせ
-
辻村真樹(M2)が令和3年度学生表彰「東京大学総長賞」および「工学系研究科長賞」を受賞しました。(2022.3.4)
2022.03.04 受賞
-
辻村真樹(M2)が、第59回生物物理学会年会にて学生発表賞を受賞しました。(2021.12.7)
2021.12.07 辻村真樹受賞
-
菅生 優(M1)がCSJ化学フェスタ2021で優秀ポスター発表賞を受賞しました。(2021.12.2)
2021.12.03 受賞
-
辻村真樹(M2)がCSJ化学フェスタ2021で優秀ポスター発表賞を受賞しました。(2021.12.2)
2021.12.03 辻村真樹受賞
-
辻村真樹(M2)が、第47回生体分子科学討論会にて優秀ポスター賞を受賞しました。(2021.6.5)
2021.06.05 辻村真樹受賞お知らせ




-
プレスリリース「光合成反応における光損傷と修復のメカニズム解明 ―傷ついたタンパク質を見つけて分解するしくみを明らかに―」(2023.11.21)
2023.11.21 プレスリリース -
プレスリリース「植物の光合成初期過程の酸素発生活性を向上させるアミノ酸変異を発見—光合成・人工光合成の光エネルギー変換効率の向上へ期待—」(2022.8.18)
2022.08.18 プレスリリース -
プレスリリース「精密分子設計による高感度MRI分子プローブの開発 〜早期診断に向けたがん関連酵素活性の生体内リアルタイム計測〜」(2022.3.31)
2022.03.31 プレスリリース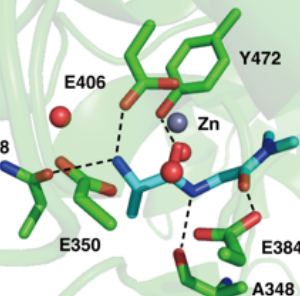
-
プレスリリース 「神経抑制に役立つタンパク質の巧みな光応答メカニズムを解明」(2021.12.21)
2021.12.21 辻村真樹プレスリリース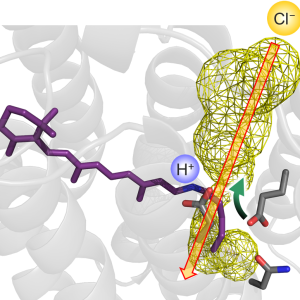
-
プレスリリース「イネの安定多収に欠かせないケイ酸チャネルの構造基盤を解明」(2021.10.29)
2021.10.29 プレスリリース
-
プレスリリース 「神経活動を操作する光スイッチタンパク質の開発」(2020.7.22)
2020.07.22 辻村真樹プレスリリース
-
天然のダイオード蛋白質
2017.04.28 プレスリリース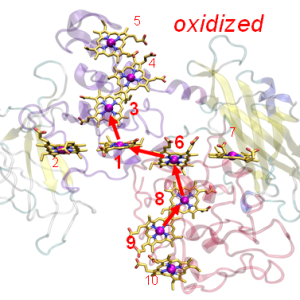
-
光合成水分解反応初期に利用される水素イオン移動経路を解明
2015.01.17 プレスリリース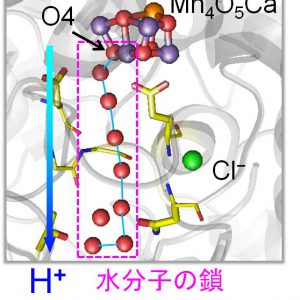
-
光合成(朝倉書店)
2021.12.13 書籍情報石北 央
-
光合成のエネルギー変換と物質変換―人工光合成をめざして(化学同人)
2015.05.09 書籍情報石北 央
-
新メンバー向けの書籍
2021.01.16 石北研について書籍情報
-
私が量子コンピュータに期待すること(東京化学同人)
2020.05.18 書籍情報石北 央
-
23の先端事例がつなぐ計算科学のフロンティア(近代科学社)
2020.01.25 書籍情報石北 央
-
ブレイクスルーへの思考: 東大先端研が実践する発想のマネジメント(東京大学出版会)
2016.12.28 書籍情報石北 央
-
酵素の振る舞いを計算で解く「2013年ノーベル化学賞を受賞したA. ウォーシェル博士に聞く」(東京化学同人)
2014.07.18 書籍情報石北 央
-
高校生・受験生が東大をもっと知るためのサイト 「キミの東大」に辻村真樹(D1)が掲載されました。(2022.5.31)
2022.06.10 辻村真樹受賞お知らせ -
「駒場生のための進学ガイダンス」にて辻村真樹(D1)が講演をしました 。(2022.4.28)
2022.04.28 辻村真樹お知らせ
-
辻村真樹(D1)が学振特別研究員DC1に採用されました。(2022.4.1)[2021.9.27内定]
2022.04.01 辻村真樹お知らせ -
辻村真樹(M2)が、第47回生体分子科学討論会にて優秀ポスター賞を受賞しました。(2021.6.5)
2021.06.05 辻村真樹受賞お知らせ -
菅生 優(B4)が、卒業論文で優秀発表賞(工学部応用化学科)を受賞しました。(2021.2.10)
2021.02.11 受賞お知らせ -
西尾 俊哉(B4)が、卒業論文で優秀発表賞(工学部応用化学科)を受賞しました。(2021.2.10)
2021.02.11 受賞お知らせ -
三橋孝司(M1)が、第58回生物物理学会年会にて学生発表賞を受賞しました。(2020.9.24)
2020.09.24 受賞お知らせ
-
辻村真樹(M1)が、第58回生物物理学会年会にて学生発表賞を受賞しました。(2020.9.24)
2020.09.24 受賞お知らせ



-
光合成という夢のある反応 (西尾 俊哉 [M2])
2022.11.11 エッセイ -
理論によって放たれる光 (神田 知樹 (M2))
2022.06.10 エッセイ -
たくさんの分野の間に(三橋 孝司 (M2))
2021.09.27 エッセイ -
理論と実験の協奏(辻村 真樹 (M2))
2021.09.17 辻村真樹エッセイ -
理論と実験、溝と橋渡し(野地 智康)
2021.06.04 エッセイ野地 智康
-
在宅ワークについて
2020.09.08 エッセイ個人ブログ斉藤 圭亮
-
温故知新
2020.05.08 エッセイ個人ブログ石北 央
-
似て非なるもの
2019.10.04 エッセイ個人ブログ石北 央
-
高校生・受験生が東大をもっと知るためのサイト 「キミの東大」に辻村真樹(D1)が掲載されました。(2022.5.31)
2022.06.10 辻村真樹受賞お知らせ -
「駒場生のための進学ガイダンス」にて辻村真樹(D1)が講演をしました 。(2022.4.28)
2022.04.28 辻村真樹お知らせ -
辻村真樹(D1)が学振特別研究員DC1に採用されました。(2022.4.1)[2021.9.27内定]
2022.04.01 辻村真樹お知らせ -
プレスリリース 「神経抑制に役立つタンパク質の巧みな光応答メカニズムを解明」(2021.12.21)
2021.12.21 辻村真樹プレスリリース -
光合成(朝倉書店)
2021.12.13 書籍情報石北 央 -
辻村真樹(M2)が、第59回生物物理学会年会にて学生発表賞を受賞しました。(2021.12.7)
2021.12.07 辻村真樹受賞 -
辻村真樹(M2)がCSJ化学フェスタ2021で優秀ポスター発表賞を受賞しました。(2021.12.2)
2021.12.03 辻村真樹受賞 -
理論と実験の協奏(辻村 真樹 (M2))
2021.09.17 辻村真樹エッセイ
-
プロトン移動は吸収波長をどのように変える?
2022.04.21 光合成・光受容プロトン移動光励起pKa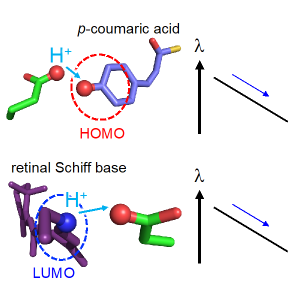
-
蛋白質中で存在が確認された低障壁水素結合の例:プロトン移動を仲介
2021.02.23 光合成・光受容電子移動プロトン移動 -
一般的な水素結合でも短くすれば低障壁水素結合性を帯びるのか?
2021.02.13 光合成・光受容酵素・創薬系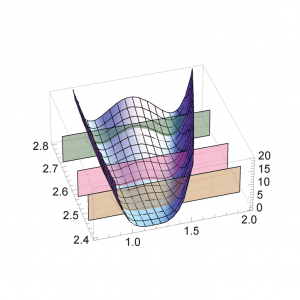
-
疎水性残基の影響を軽視しない:旧式の理論化学研究が陥る罠
2021.02.13 光合成・光受容酵素反応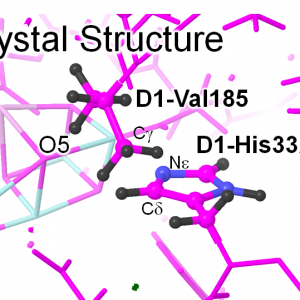
-
「低障壁水素結合」の判定
2021.02.13 プロトン移動酵素・創薬系pKa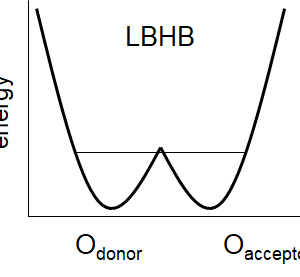
-
蛋白質中のクロロフィルの吸収波長はどうして変化する?
2021.01.24 光合成・光受容光励起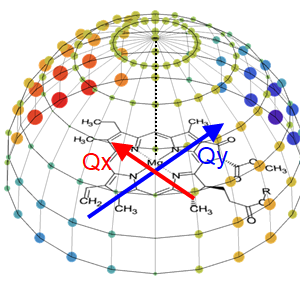
-
「結晶構造座標=実験結果」と鵜呑みにしない:理論から訂正した「結晶構造」
2021.01.18 電子移動酵素・創薬系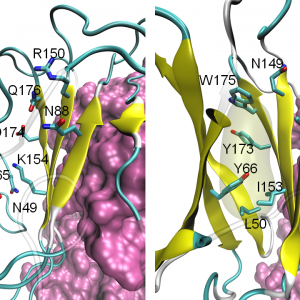
-
微生物型ロドプシンの吸収波長を決める本質的な要因
2021.01.16 プロトン移動光励起酵素反応
-
プロトン移動は吸収波長をどのように変える?
2022.04.21 光合成・光受容プロトン移動光励起pKa -
蛋白質中で存在が確認された低障壁水素結合の例:プロトン移動を仲介
2021.02.23 光合成・光受容電子移動プロトン移動 -
一般的な水素結合でも短くすれば低障壁水素結合性を帯びるのか?
2021.02.13 光合成・光受容酵素・創薬系 -
疎水性残基の影響を軽視しない:旧式の理論化学研究が陥る罠
2021.02.13 光合成・光受容酵素反応 -
「低障壁水素結合」の判定
2021.02.13 プロトン移動酵素・創薬系pKa -
蛋白質中のクロロフィルの吸収波長はどうして変化する?
2021.01.24 光合成・光受容光励起 -
「結晶構造座標=実験結果」と鵜呑みにしない:理論から訂正した「結晶構造」
2021.01.18 電子移動酵素・創薬系 -
微生物型ロドプシンの吸収波長を決める本質的な要因
2021.01.16 プロトン移動光励起酵素反応
-
一般的な水素結合でも短くすれば低障壁水素結合性を帯びるのか?
2021.02.13 光合成・光受容酵素・創薬系 -
疎水性残基の影響を軽視しない:旧式の理論化学研究が陥る罠
2021.02.13 光合成・光受容酵素反応 -
「低障壁水素結合」の判定
2021.02.13 プロトン移動酵素・創薬系pKa -
「結晶構造座標=実験結果」と鵜呑みにしない:理論から訂正した「結晶構造」
2021.01.18 電子移動酵素・創薬系 -
微生物型ロドプシンの吸収波長を決める本質的な要因
2021.01.16 プロトン移動光励起酵素反応 -
X線自由電子レーザーによるS3状態構造のO…O = 1.9Åの化学種とは?
2021.01.16 電子移動プロトン移動酵素反応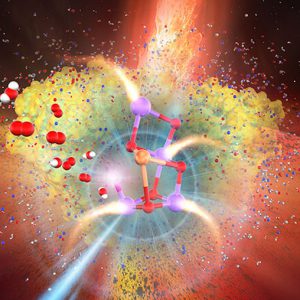
-
Mn…Oの長さでradiation-damageを語ってよいものか?
2021.01.16 電子移動酵素反応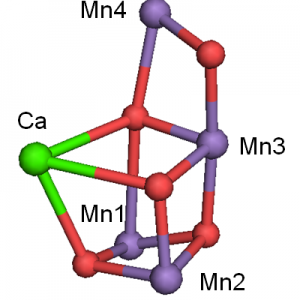
-
水分解酸素発生反応におけるCa2+の役目は?
2021.01.16 電子移動プロトン移動酵素反応
-
プロトン移動は吸収波長をどのように変える?
2022.04.21 光合成・光受容プロトン移動光励起pKa -
「低障壁水素結合」の判定
2021.02.13 プロトン移動酵素・創薬系pKa -
低障壁水素結合の酸化還元電位:H+位置依存性
2021.01.16 光合成・光受容電子移動プロトン移動pKa電位
-
実験では測定不可能なキノン分子のpKaを電子状態から求める
2021.01.16 光合成・光受容プロトン移動新しい理論化学手法の開発pKa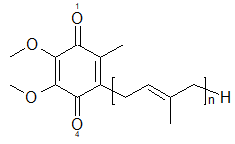
-
蛋白質中のプロトン移動経路・水分子鎖のpKaを求める
2021.01.16 光合成・光受容プロトン移動新しい理論化学手法の開発pKa
-
ジャーナル・カバーアート・ギャラリー
2021.01.14 光合成・光受容酵素・創薬系新しい理論化学手法の開発
-
光合成におけるプロトン移動
2020.12.28 光合成・光受容プロトン移動新しい理論化学手法の開発QM/MM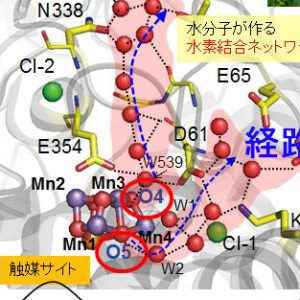
-
光合成における電子移動
2020.12.26 光合成・光受容電子移動新しい理論化学手法の開発電位