
蛋白質の活性部位を理論化学的手法で研究しようとする際に、「重要な部位(例:酵素の触媒活性部位)だけを蛋白質の結晶構造から切り出して計算を行う」。そういった研究が以前は多く見られました。現在は、蛋白質分子を取り扱う際はQM/MM法 (quantum mechanical/ molecular mechanical approach) を利用する機会が増えているため、そういった研究手法で発表された論文は減りつつありますが、それでもまだ見受けられます。
「重要な部位」を人が主観で判断すると…
そもそも蛋白質の研究を行う際に、「重要な部位」を人はどうやって見分けることができるのでしょうか。そこが問題です。その人の主観が入ったり、あるいは人的ミスが誘発される余地があります。
例えば以下のような例があります。photosystem II (PSII)における最も低酸化状態のMn4CaO5であるS0状態が、一電子酸化によりS1状態になる過程「S0→S1遷移」について、よく引用されているSiegbahnの研究です。PSII結晶構造ではO5周りのMn…O5距離が長めであるため、結晶構造を見て誰もが(計算せずとも)「O5が基質水分子のOではないか」と推察したくなります。そういった仮説を立て、実証を行い、うまくいけばその結論でも構いません。ただ、研究者も所詮人であり、気をつけないと先入観や欲望でサイエンスと思った結果が、実はねじ曲がったものであるかもしれません。
Siegbahnは「O5…Mnが長いのはO5がOH-であるから」とし、「O5が基質の候補」と仮説を立てました。基質の水分子(=酸素となる水分子)であればH+が外れていく必要があるためO5のOH-も解離する必要、すなわちH+移動(H+放出)する必要があります。H+移動のためにはH+アクセプターとなる水素結合パートナーが必要です(注意:ここは蛋白質内部であり、バクル領域ではない)。しかし、結晶構造においてO5の周辺には水素結合できる水分子は存在しません。そう考えればO5からのH+は起こりにくいと考えるのが、基礎の化学を理解している人なら素直に至る、自然な考え方です。しかし、Siegbahnは「S0→S1の過程において、結晶構造では見えない(動きやすい)水分子がO5に近寄り、O5のOH-がこの水分子にH+を渡すことでH+放出は可能」とし、プロトン放出過程のエナジェティクスを以下のような構造に基づいて彼なりに「実証」しました。
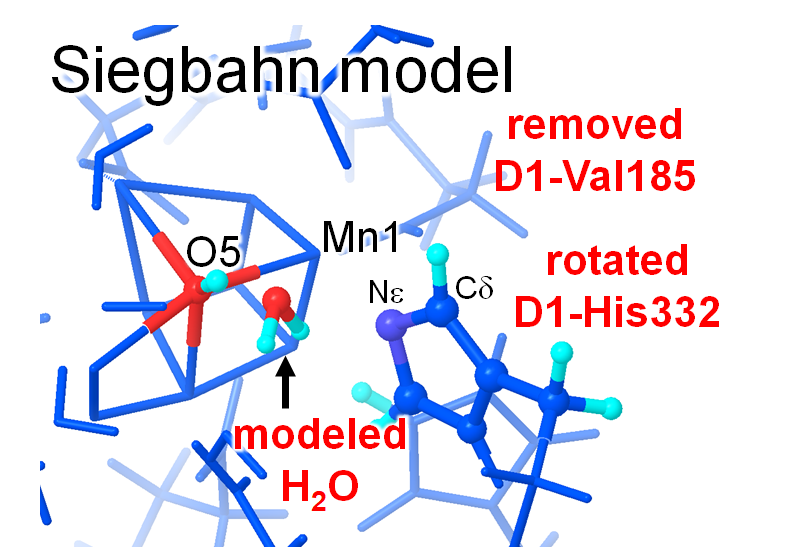
図 SiegbahnのS0状態のモデル
(自ら手を動かして本当の結晶構造を考察せず)彼の論文中の図だけを見て論文を読み進めると、「なるほど」と思われるかもしれません。マニアックな考え方をよく思いついたな、とも思います。しかし、実際の結晶構造は以下のようになっています。両者をにらめっこして、間違い探しをしてみてください。
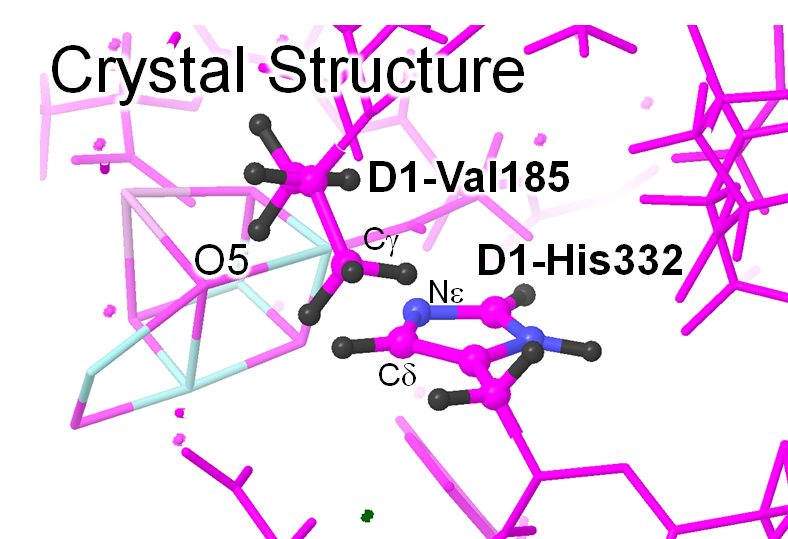
図 PSII結晶構造 (PDB ID: 3ARC)
間違い探しはできましたか? 答えは以下:
(1) Siegbahnが水分子をおいたO5近傍の位置には、現実のタンパク質結晶構造では疎水性アミノ酸残基D1-Val185が存在する。
→疎水的環境に水分子は好んでこないし、そもそもVal185側鎖の立体障害で水分子座標を置くことは無理。
(2) Mnの配位子であるD1-His332側鎖の配向が90度異なる。
→実は、この操作もあわせて行わないと、彼がモデルした水分子はHis配位子と立体障害を起こす。ちなみに、結晶構造ではHis残基は周辺のアミノ酸残基と水素結合をしており、90度回転は許されません。
答えは以上です。
得られる教訓
「蛋白質分子をまるごと扱うのは大変だから重要な部位を切り出す」作戦に長年慣れてしまっていると、以下の作業を特に気にせず平然と行ってしまうのかもしれません:
(1) 疎水性残基との静電相互作用は無視できるから、活性にとって重要ではない。削除しても、周囲の研究者からも理解が得られるだろう。
→極性が高い水分子の存在を意識していれば、水分子のアクセスが制限できる蛋白質内部の環境においてそのような「操作」はしてはいけません。疎水性残基の側鎖を削除することは、ありもしないcavityを人為的に作り出すこと。現実のタンパク質では、一般的な量子化学計算のような「cavity=何もない真空」ではありません。cavityができれば、現実では極性の高い水分子が侵入することになります。つまり、Siegbahnの操作はV185G等のアミノ酸変異を作り出しているのと同等な作業であり、計算結果は少なくともwild typeのものには対応しないと思います。
(2) 配位子の回転くらい許されるだろう・大目に見てくれ
→小分子の錯体等でバルクに直接配位子が露出しているようなケースでは、配位子の回転は許されるケースもあると思います。しかし、蛋白質の内部、特にPSIIのこの領域はスカスカではないです。
これらの問題は、QM/MMで取り扱っていれば全て防げた(モデルとして捏造になるためあり得なかった)ことだったと思います。QM/MM計算では、たとえ研究を行った人の目が節穴だったとしても、D1-Val185は少なくともMM領域として考慮されるため、その側鎖の体積により水分子がO5近傍に入る余地はありません。
ということで、O5基質説としてS0→S1過程におけるH+放出をどうしても語りたい際は、上記の問題を熟知した上、自身の胸に手を当てた上でそれでも良心がとがめないか熟慮の上で行う覚悟が必要です。(というか、そこまでして、わざわざ語りたいですか?そう言う人は、科学者としての良心はどこへ行ってしまったのか、考えた方がいいかも。)
以上の問題は、石北研究室の研究で初めて丁寧にSiegbahnの座標から解析を行うことで明らかになりました。(みんな、論文に書いてあることをそのまま信じてしまうようです。Supporting Informationなど飾りに過ぎないのかな、と思います。)権威がある人がみんなの耳に都合の良い説(例:O5からのH+放出機構)を主張すると、人は批判せずに受け入れがちです。O5は特殊な位置にあるため、もし本当に「O5が基質でS0→S1でH+が放出」ならば私たちも本当に気が楽なのですが。。H+放出を説明できるモデルが現状ではないのです。この過程もきちんと説明できるのであればO5基質はかなり有力となります。少なくともサイエンスにおいては、イソップ物語の裸の王様にでてくる(忖度するような大勢の取り巻きの大人ではなく)子供でありたいですね。
石北研究室の以下の論文でまとめてありますので引用してご活用ください:
Keisuke Saito, A. William Rutherford, and Hiroshi Ishikita*
Nat. Commun. 6:8488 (2015) doi: 10.1038/ncomms9488
“Energetics of proton release on the first oxidation step in the water-oxidizing enzyme”
Journal Pubmed
Hiroshi Ishikita*
Oxygen Production and Reduction in Artificial and Natural Systems (James Barber, Alexander V Ruban, and Peter J Nixon, Eds.) (2019) pp 191-208, World Scientific, doi: 10.1142/9789813276925_0010, ISBN: 978-981-3276-93-2
“Protein Environment that Facilitates Proton Transfer and Electron Transfer in Photosystem II”