
Ca2+はphotosystem IIの水分解酸素発生触媒部位Mn4CaO5錯体に組み込まれています。Ca2+そ欠損した錯体でも、光を一回照射せたS1→S2遷移は問題なく起こります。一方、計2回照射させたS2→S3遷移は阻害されてしまいます。だからCa2+は水分解反応に必須のcomponentであることは明らかです。一方で、なぜ反応が阻害されるか=なぜ必須なcomponentなのか、というメカニズムに関してはわかっていませんでした。
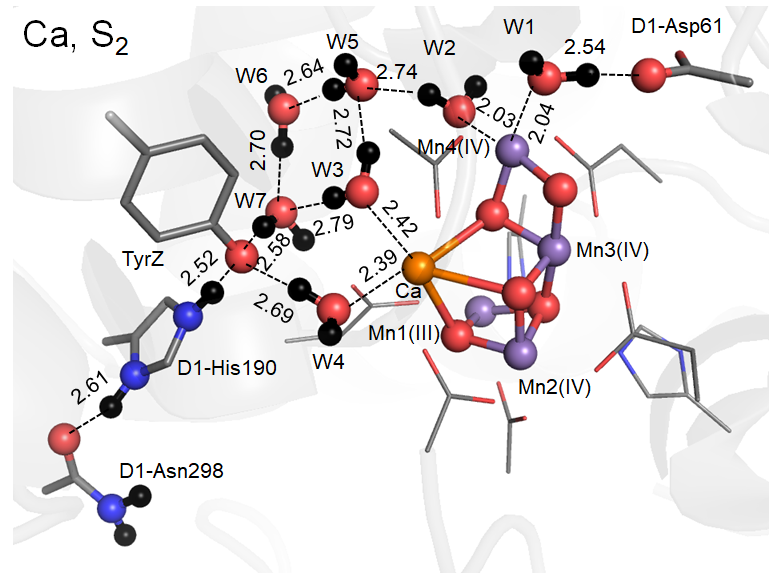
Ca2+はMn4CaO5錯体の配位水分子4つのうち2つ(W3,、W4)の結合サイトを提供しています。W3、W4はdiamond-shaped clusterと呼ばれる水分子クラスターによる水素結合ネットワークでMn4CaO5の電子アクセプターとなるTyrZ…D1-His190の低障壁水素結合ペアともつながっています。
S2→S3遷移では、TyrZへの電子移動に伴うMn4CaO5錯体の1電子酸化のみならず、Mn4CaO5錯体からの1H+放出も伴います。さらに上述の分子構造から、Ca2+欠損による水分解反応の阻害として(1)プロトン移動の阻害、(2)電子移動の阻害、の可能性があると長い間議論されていました。
プロトン移動
S2→S3遷移ではプロトン移動はdangling Mn4に配位している配位水W1から水素結合アクセプターであるD1-Asp61が低障壁水素結合を形成することで起こります(論文)。
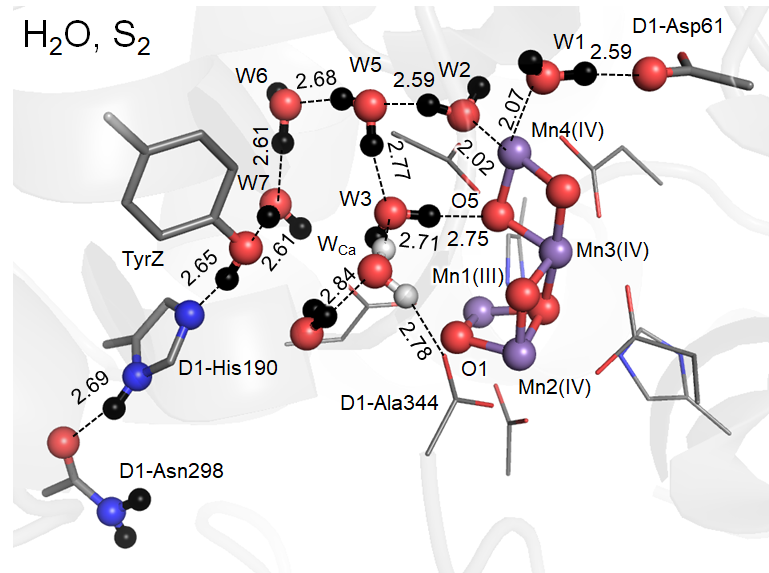
Ca2+欠損構造ではCa2+欠損部位をH2O・低pHではH3O+が占めますが、W1…D1-Asp61間の水素結合はもはや低障壁水素結合ではなく、H+はW1に存在しD1-Asp61へはuphillのH+移動となる通常の水素結合となってしまうことが石北研究室により解明されました。その理由は、Ca2+の+2が消失するため、W1のpKaが上昇しプロトンを放出しにくくなるためです。すなわり、H+移動も阻害されることが分子化学的に初めて示されました。
電子移動
近年開発した手法で、石北研究室では世界で初めてphotosystem II水分解電子移動経路の酸化還元電位を全て明らかにしました(論文)。そして、native photosystem IIでは確かにMn4CaO5 → TyrZ…D1-His190 → P680と電子移動カスケードを作っている様を明らかにしました。
Ca2+が欠損するとMn4CaO5 → TyrZ…D1-His190の電子移動がuphillになる事実が石北研究室によって初めて明らかになりました。しかし、この結果をあなたはありのまま受け入れられるでしょうか?違和感を感じた人は化学をしっかり理解している人だと思います。
Ca2+という+2がなくなったのなら、Mn4CaO5の電位は負にシフトし、逆に電子ドナーとして電子を放出しやすくなり電子移動は起こりやすくなる、とまず考えるのが筋です。
実は、石北研究室によって明らかになった電位でも、Ca2+欠損によりMn4CaO5の電位は確かに負にシフトしています。しかし、それ以上に、電子アクセプターであるTyrZ…D1-His190の電位が府側にシフトするため、トータルとしてMn4CaO5 → TyrZ…D1-His190の電子移動がuphillになるのです。
つまり、Mn4CaO5のcomponentであるCa2+は、己の属しているMn4CaO5の酸化還元電位以上に、5 Å程度はなれたTyrZ…D1-His190の電位により大きな影響を与えているのです。面白いと思いませんか?
でもなぜそうなるのでしょうか。その理由は、上述の通りCa2+にはW3、W4の二つの配位水があるため、水分子のOがMn4CaO5に向いている配向となっています。ここでCa2+欠損により水分子の配向が大きく変わります。そもそもカチオンであるCa2+をMn4O5骨格が保持するために、結合サイトとして3つの酸素原子(O1, O2, O5)を利用しているため、Ca2+がなくなると、今度は水分子のHがMn4O5側に向いてきます。この効果は、Mn4CaO5にとってはむしろ消失したCa2+の正電荷を補うため電位の低下を緩和してくれるのですが、その先の水素結合ネットワークにあるTyrZ…D1-His190には、水素結合ネットワークの配向を大きく換えてしまうこともあって、W3、W4の配向変化以上のdipoleの変化を伴い、電位がより大きくシフトしてしまうことになります。
こういった、単分子のケミストリーでは捉えきれないネットワークを介した反応機構が、蛋白質反応場では起こるわけであり、これが面白いところです。また、ここまで解き明かすことは、実験だけしていたり、結晶構造を眺めていたりするだけではなかなか難しいと思います。理論と実験が車軸の両輪として機能していくことが、サイエンスのためにもハッピーだと思います。
以上は本来複雑な現象ですが、石北研究室の研究により、化学の基礎がある学部生にも十分理解できる平易な言葉でその制御機構が明らかににされました。以下の論文に記載されています。引用してご活用ください。
Keisuke Saito, Manoj Mandal, and Hiroshi Ishikita*
Biochemistry 59 (2020) 3216-3224. doi: 10.1021/acs.biochem.0c00177
“Energetics of ionized water molecules in the H-bond network near the Ca2+ and Cl– binding sites in photosystem II”
Journal Pubmed